 Identificaciones de bdstaylor
Identificaciones de bdstaylor




Fotos / Sonidos

Observ.
manuel_helsinkiDescripción
In a water sample from a bay/marshland in the Baltic Sea.
 peptolab, algunos derechos reservados (CC BY-NC)")
 peptolab, algunos derechos reservados (CC BY-NC)")
Fotos / Sonidos

Observ.
dgborinDescripción
Sample taken on 2024-07-20.
Slowing solution 0.5% Methyl cellulose.
 Igor Shelpiakov, algunos derechos reservados (CC BY), subido por Igor Shelpiakov")
Fotos / Sonidos

Observ.
closterium_mysteriumDescripción
Video: https://youtu.be/lSif6qvuMEQ
Sampling location: A sand sample was collected from Adzghapsha Beach, located on the coast of the Black Sea. The sand was almost dry at the time of collection.
Date and time of collection: September 9, 2023 at 11 AM
The sample was stored in a sealed glass container at room temperature.
In about a week, a small amount of this sand was mixed with
saltwater prepared from distilled water and marine aquarium salt to achieve a salinity of approximately 20 g/kg. However, determining the precise salinity level is impossible without a salt meter.
Date and time of observation: September 22, 2023 at 11 AM
All 100x objective images and videos are of the same organism.
All 10x and 40x objective images and videos are of a few other ciliates from the same water sample.
Diagnosis of the organisms observed:
• Slender, flexible cell
• Three longitudinal cortical grooves that allow the cell to spiral the anterior half of the cell
• Spiral movement when swimming
• Length 60-75 µm
• Colorless cortical granules widely distribute in the cortex
• Buccal opening approximately one third of total length
• A clear subequatorial CV. At least two other small vacuoles were visible in line with the large CV at the posterior part of the cell
Unfortunately, I am not sure about the other important features, such as the nuclear apparatus and the arrangement of the kineties. The tendency of this ciliate to bend its body in a spiral and my lack of experience confused me.

Fotos / Sonidos

Observ.
douchDescripción
In scum at exit of drainage pipe. Bright-field light microscopy.
 Karolina Fucikova, algunos derechos reservados (CC BY-NC), subido por Karolina Fucikova")
Fotos / Sonidos
Qué
Género TrinemaObserv.
douchDescripción
In scum at exit of drainage pipe. Bright-field light microscopy.
 Vicente Franch Meneu, algunos derechos reservados (CC BY-NC), subido por Vicente Franch Meneu")
Fotos / Sonidos

Observ.
douchDescripción
In scum at exit of drainage pipe. Bright-field light microscopy.
Fotos / Sonidos

Observ.
douchDescripción
In scum at exit of drainage pipe. Bright-field light microscopy.
Image 1: central focal-plane
Image 2: upper focal-plane
Image 3: lower focal-plane
 crseaquist, algunos derechos reservados (CC BY-NC), subido por crseaquist")
Fotos / Sonidos

Qué
Orden SessilidaObserv.
douchDescripción
In scum at exit of drainage pipe. Bright-field light microscopy.
Image 1: central focal-plane
Image 2: upper focal-plane
Image 3: lower focal-plane

Frank Fox, algunos derechos reservados (CC BY-SA)")
Fotos / Sonidos

Qué
Género CothurniaObserv.
douchDescripción
In scum at exit of drainage pipe. Bright-field light microscopy.
Image 1: upper focal-plane
Image 2: central focal-plane
Image 3: lower focal-plane
Image 4: contracted down test
 peptolab, algunos derechos reservados (CC BY-NC)")
 Proyecto Agua, algunos derechos reservados (CC BY-NC-SA)")

Fotos / Sonidos

Observ.
douchDescripción
In algae adhered to intertidal rock. Bright-field light microscopy.
 Vicente Franch Meneu, algunos derechos reservados (CC BY-NC), subido por Vicente Franch Meneu")

 Vicente Franch Meneu, algunos derechos reservados (CC BY-NC), subido por Vicente Franch Meneu")
Fotos / Sonidos

Qué
Trachelius ovumObserv.
davidfbirdDescripción
Eutrophic reservoir. The cytostome is visible as a funnel structure at the base of the neck. 330 µm long.
 cwwood, algunos derechos reservados (CC BY-SA), subido por cwwood")

Fotos / Sonidos

Observ.
someplantDescripción
Magnification of photos: 100×, 100×, 100×, 100×, 200×, 200×, 200×, 200×, 400×, 400×, 400×, 400×, 400×, 400×, 400×, 400×, 600×
Habitat: Saltwater squeezed out of beach mud (by stepping on it), near a saltmarsh. I presume the mud was hypoxic due to its rotten-egg smell.
Photo taken with a Celestron PentaView Digital Microscope. According to their website, the FOV (i.e. the diagonal width) at 100× is 600 µm.
Videos available: https://youtu.be/FIey5h5ooyg
https://youtu.be/ifWMmM7w1us
 peptolab, algunos derechos reservados (CC BY-NC)")
Fotos / Sonidos

Observ.
someplantDescripción
Magnification of photos: 100×, 100×, 100×, 200×, 200×
Habitat: Saltwater squeezed out of beach mud (by stepping on it), near a saltmarsh. I presume the mud was hypoxic due to its rotten-egg smell.
Photo taken with a Celestron PentaView Digital Microscope. According to their website, the FOV (i.e. the diagonal width) at 100× is 600 µm.
Video available: https://youtu.be/AoxdmozIkyM
 Ashish Aswin Kumar, todos los derechos reservados, subido por Ashish Aswin Kumar")

 Proyecto Agua, algunos derechos reservados (CC BY-NC-SA)")

 Bruce Taylor, algunos derechos reservados (CC BY), subido por Bruce Taylor")
Fotos / Sonidos
Observ.
rqy-yongDescripción
Tail a long, anchored coiled spring which would gradually extend then snap back instantly
 Proyecto Agua, algunos derechos reservados (CC BY-NC-SA)")
Fotos / Sonidos
Observ.
amayakanDescripción
Video: https://youtu.be/0ZX_yjqOjk8
Cell size: 33-40 µm in length
Site of sample collection: Takamori Chuo Park (a freshwater habitat), Sendai, Japan
Date of sample collection: June 18th, 2022
Weather: Sunny
Water temp.: 22.3°C
pH 6.6
Date of observation: June 18th, 2022
Bright field observation using a Wraymer microscope (model BX-3500TL, Osaka, Japan) equipped with a Floyd-2 HDMI ethernet digital camera (Wraymer, Osaka, Japan). The accuracy of the scale bar was confirmed by using a stage micrometer glass slide (1 div. = 10 µm; Wraymer, Osaka, Japan) at each magnification.
 Picturepest, algunos derechos reservados (CC BY)")
Fotos / Sonidos

Qué
Género GruberiaObserv.
someplantDescripción
Magnification of photos: 100×, 200×, 200×, 200×, 200×, 400×, 400×, 400×, 400×, 400×, 600×, 600×, 600×, 100×, 200×, 100×, 200×, 200×, 400×, 100×
Habitat: Saltwater squeezed out of beach mud (by stepping on it), near a saltmarsh. I presume the mud was hypoxic due to its rotten-egg smell.
Photo taken with a Celestron PentaView Digital Microscope. According to their website, the FOV (i.e. the diagonal width) at 100× is 600 µm.
Video available: https://youtu.be/9ug8vwRZt3U
Several individuals present in sample.
 Phaulactis, algunos derechos reservados (CC BY-NC), subido por Phaulactis")
Fotos / Sonidos

Qué
Lacrymaria olorObserv.
hgmuellerDescripción
Found another, this time showing the macronuclei
 Picturepest, algunos derechos reservados (CC BY)")
Fotos / Sonidos
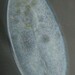
Qué
Frontonia leucasObserv.
hgmuellerDescripción
L=260 um, W=120 um Lmouth=34 um
Oral apparatus 1/8 body length, arch shaped and near end of 1/4 body. A little doubtfull because I couldnt find the radiating water canals on the CV.
")

Estadísticas
- 7859



























