Does the colouration of the giant panda hint of anti-predator defence by means of mutilating premolars? part 1
(For a concise version please see https://www.inaturalist.org/journal/milewski/66782-why-is-the-giant-panda-black-and-white-concise-version#)
Why is the giant panda (Ailuropoda melanoleuca, https://www.inaturalist.org/taxa/41659-Ailuropoda-melanoleuca) so boldly black and white?
Why - in contrast to other bears - is its colouration so consistent, i.e. lacking individual variation?
And why does it tend to be so unexpressive?
(The main function of the checker-like pattern of the giant panda is unlikely to be camouflage in a patchily snow-covered landscape, as a few authors have previously suggested.)
Could this plausibly be an example of warning colouration?
Is it even possible that the giant panda is the largest-bodied carnivore that possesses aposematic colouration at the scale of the whole body?
The most dangerous predators - apart from humans - historically affecting the giant panda were
- the tiger (Panthera tigris amoyensis, https://www.inaturalist.org/taxa/41967-Panthera-tigris) and
- the dhole (Cuon alpinus hesperius/lepturus, https://www.inaturalist.org/taxa/42101-Cuon-alpinus).
Also playing a part were
- the leopard (Panthera pardus delacouri/japonensis, https://www.inaturalist.org/taxa/41963-Panthera-pardus) and
- the snow leopard (Panthera uncia uncioides, https://www.inaturalist.org/taxa/74831-Panthera-uncia).
The giant panda was in all likelihood vulnerable to predation because of its noisy eating habits and relatively slow reproduction.
All members of the Carnivora defend themselves with their teeth, and many show their teeth to would-be attackers.
However, the giant panda seldom adopts a fang-baring expression or even opens its mouth in fear/anger.
Furthermore,
- the canines are relatively small (https://www.shutterstock.com/nb/image-photo/panda-sits-growls-turning-her-head-1560141806 and https://www.quora.com/Are-pandas-aggressive-or-calm) and would show poorly against the whitish muzzle eve if such an expression was adopted, and
- the claws are short and unimpressive compared with other bears, particularly the sun bear (Helarctos malayanus, https://www.inaturalist.org/taxa/41655-Helarctos-malayanus), which has extremely dangerous claws according to injured zookeepers.
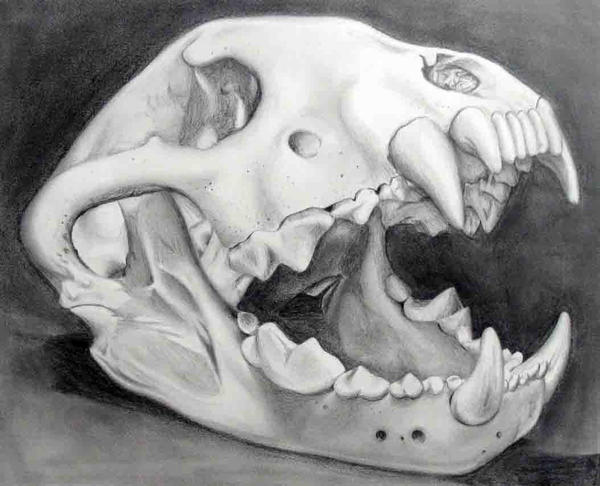
What seems to have been overlooked about the giant panda is that its premolars are unique among carnivores in their shape and arrangement (https://animaldiversity.org/accounts/Vertebrata/specimens/collections/contributors/skulls/ailuropoda/a._melanoleuca/panda_ltr/?f=subject::specimen::teeth::lower%20tooth%20row and https://commons.wikimedia.org/wiki/File:Animaldentition_ailuropodamelanoleuca.png).
Also see https://upload.wikimedia.org/wikipedia/commons/4/45/Giant_Panda_Skull.JPG and https://boneclones.com/product/giant-panda-skull-adult-BC-324 and https://www.skeletonsandskullssuperstore.com/product/giant-panda-female-skull/). These premolars may not look intimidating (https://www.alamy.com/closeup-of-giant-panda-s-mouth-and-teeth-wolong-nature-reserve-sichuan-image106119.html), but would-be predators would be wise to beware.
The giant panda has the most powerful bite of any extant species of bear (https://www.researchgate.net/publication/224630206_Finite_Element_Analysis_of_ursid_cranial_mechanics_and_the_prediction_of_feeding_behaviour_in_the_extinct_giant_Agriotherium_africanum_the_bare_facts and https://zslpublications.onlinelibrary.wiley.com/doi/full/10.1111/j.1469-7998.2011.00862.x). The bite is powerful even at the premolars, because
- the jaws are short and the cheek-teeth are crowded right behind the canines, and
- the masseter muscles are extremely developed, in conjunction with a high sagittal crest (https://en.wikipedia.org/wiki/Sagittal_crest) and wide zygomatic arches (https://en.wikipedia.org/wiki/Zygomatic_arch).
See https://phys.org/news/2021-07-evolution-giant-panda-temporomandibular-joint.html and https://news.cgtn.com/news/3d3d674d7763444d32457a6333566d54/index.html and https://www.researchgate.net/figure/Adaptive-morphological-characteristics-of-giant-pandas-A-pseudothumb-denoted-by-the_fig1_266570673.
The giant panda has the shortest face of all bears. Ironically, the same short muzzle that makes the giant panda rather appealing as a 'human face' also houses its main weaponry.
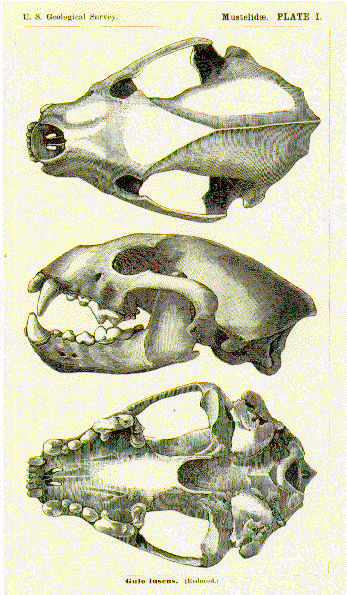
The compact dentition of the giant panda can best be illustrated in contrast with the polar bear, the species most specialised for carnivory.
Please compare https://www.mindenpictures.com/stock-photo-giant-panda-ailuropoda-melanoleuca-skull-adult-length-mm-naturephotography-image80161382.html and https://www.zoochat.com/community/media/skull-giant-panda.469645/ with https://beautifullybony.wordpress.com/archive/bone-of-the-month/the-polar-bear/the-skull-biomechanics-of-the-polar-bear/ and http://digimorph.org/specimens/Ursus_maritimus/male/ and https://www.westend61.de/en/imageView/MW000023/skull-of-polar-bear-ursus-maritimus-in-front-of-black-background-side-view and https://www.acornnaturalists.com/bear-polar-skull-replica.html.
Among living bears, it is the least carnivorous species, i.e. the giant panda, that has
- the strongest bite, and
- the largest premolars.
Bamboo, a woody form of grass, is the staple diet of the giant panda.
An adaptive reason for the powerful bite at the level of the premolars may be an ability to slice bamboo, preparatory to grinding it with the molars (https://www.researchgate.net/figure/Segmented-maxillary-teeth-of-the-living-bear-species-analysed-in-this-article_fig3_328232826 and https://www.natureinstock.com/search/preview/giant-panda-ailuropoda-melanoleuca-skull-adult-close-up-of-molar/0_11300923.html and https://animaldiversity.org/accounts/Carnivora/specimens/collections/contributors/skulls/ailuropoda/a._melanoleuca/panda_lm/).
In particular, premolars 2 and 3 on the upper jaw, and 2, 3 and 4 on the lower jaw, form a tightly occluding set, operating by means of powerful jaw-muscles. (I have described these teeth in detail in the comments below.)
Although the dentition of the giant panda may be adaptive primarily to its diet, the same premolars are 'mutilars', capable of mutilating any animal attacking the giant panda. This is in conjunction with clamping by the jaws and hugging with the forelegs - in contrast to the piercing bite-and-release and scratching inflicted by felids.
The key to understanding the individually consistent black-and-white pattern of the giant panda as warning colouration may be to realise that the bamboo-crushing premolars of this species are quasi-carnassials, and that they are
- even more dangerous than the canines of most other carnivores, but
- intrinsically difficult to display directly for the hazard that they are.
According to this explanation, the giant panda may have evolved true aposematic whole-body colouration, as a way of announcing its hidden defensive capability.
In its strict sense, 'aposematic' refers to warning colouration in those organisms in which the defensive capabilities are not only extraordinary and specialised, but also non-apparent as such.
If the giant panda really is aposematic at the scale of the whole body, then it may possibly be the largest-bodied aposematic organism on Earth.
How do the patterns of dark/pale contrast differ between the giant panda and other bears?
I will discuss the eyes and ears in part 2. However, in the case of the chest, other species of bears show certain trends in colouration that can be extrapolated to the extremes shown in the giant panda.
More than any other family of carnivores, bears express two patterns consistently:
- all bears except for the brown bear (Ursus arctos) tend to be conspicuously dark or pale overall, and
- in the dark species there tend to be conspicuously pale marking in various configurations on the chest, exposed when the animal rears up in confrontation, and possibly functioning as warning signs.
The giant panda is odd in having large patches of black on an otherwise whitish coat. Other bears tend to have either dark or pale tones dominant, without gross variegation.
Pale bears are the polar bear (Ursus maritimus) and the pale morph of the American black bear (Ursus americanus). No predominantly pale bear has dark markings on its fur.
The retention of brownish on its posterior (https://www.istockphoto.com/photo/the-giant-panda-or-bamboo-bear-gm601399942-103438177 and https://www.shutterstock.com/nb/video/clip-1021033759-giant-panda-ailuropoda-melanoleuca-rubbing-tail-against and https://www.shutterstock.com/nb/video/clip-32900209-panda-forest and https://pandathings.com/learn-about-the-giant-panda/do-pandas-have-tails/) suggests that the giant panda is not unique among bears, just extreme in its differentiation into dark versus pale.
An explanation for the relatively plain coats of other bears is that these formidable and non-gregarious mammals, all of which rely on smell and none of which hunts by stealth, need neither cryptic colouration nor social colouration that advertises sex, age, or maturity.
The dark chest of the giant panda is unusual among bears in lacking the ominous 'pectoral flags' seen in the sloth bear, Asian black bear, and sun bear, in upright confrontation (https://www.reddit.com/r/NatureIsFuckingLit/comments/ogont1/sloth_bear_stands_its_ground_against_a_tiger_india/ and https://en.wikipedia.org/wiki/Asian_black_bear#/media/File:Ursus_thibetanus_3_(Wroclaw_zoo).JPG and https://www.bigstockphoto.com/image-330810433/stock-photo-asiatic-black-bear-standing-and-relax-in-the-summer-black-bear-with-chest-the-v-shape-is-white-woo and http://www.bearsoftheworld.net/sun_bears.asp and https://www.alamy.com/stock-photo-a-sun-bear-helarctos-malayanus-standing-up-on-it-hind-legs-at-rare-89813043.html).
Why does the giant panda seems to differ categorically from other bears of similar body size in this way?
A possible interpretation is that it has effectively expanded the insignia on chest and face, to convert the whole front to black-and-white (http://www.ecns.cn/m/visual/hd/2018/01-14/151448.shtml and http://www.china.org.cn/environment/2015-04/08/content_35268385_4.htm).
Please note that, when the giant panda rears up in confrontation,
- the blackish of the chest, shoulders and forelegs contrasts sharply with the mainly whitish face and belly, and
- any blurred pattern on the hindquarters is out of view..
Thus the whole black-and-white front of the giant panda is plausibly an extension of – to the point of substituting for – the 'pectoral flags' of other bears.
to be continued in https://www.inaturalist.org/journal/milewski/66777-does-the-colouration-of-the-giant-panda-hint-of-anti-predator-defence-by-means-of-mutilating-premolars-part-2#...




























{kind=link}
{kind=link}
.JPG){kind=link}
.Is){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}